*Corresponding Author:
Vishal Srivashtav,
Department of Genetics and Plant Breeding, Rajiv Gandhi South Campus, Banaras Hindu University, Mirzapur-231001, UP, India
E-mail: vishal_bt85@yahoo.com
Abstract
Heavy metal can emanate from various natural and anthropogenic activities in various compartments viz soil, water and air around the globe. However, contamination of soil with heavy metals such as Fe, Mn, Zn, Cd and Pd have modified plant morpho-physiological growth and development which adversely affects crop production, quality and ultimately the ecosystem. Plants have developed some detoxification and tolerance strategies to cope up with this abiotic stress factor which includes accumulation and translocation, extracellular avoidance, intracellular detoxification pathways (metal sequestration, chelation and activation of antioxidant defence mechanism), signalling pathways and genetic engineering. There are various tolerance mechanisms of plants for heavy metal stress tolerant covering the role of transporters, phytochelatins, metallothioneins, organic and amino acids, proline as well as the role of phytohormones. Amongst phytohormones melatonin a ubiquitous molecule plays an important role to antioxidant activity, also as an important redox regulator, thus it is defined as bio-stimulatory of plant growth under biotic and abiotic stress. Various studies have noted that environmental stress can boost the level of endogenous melatonin in plants and exogenous application of melatonin can enhance the capability of plants to tolerate abiotic stresses. This paper presents a comprehensive review of recent approaches towards the development of phytoremediation potential using melatonin and genetic engineering with plant transformation and implementing current achievements in chloroplast genome transformation, nucleus transformation and gene stacking. Furthermore, genetic engineering of heavy metal-responsive genes has shown surprising results but its full potential remains to be exploited.
Keywords
Genetic engineering; Heavy metals; Phytohormones; Phytoremediation; Signal transduction
Abbreviations
ABC-ATP binding cassette
PCs-Phytochelatins
AMF-Arbuscular Mycorrhizal Fung
CaCA-Cation Antiporter
CDF-Cation Diffusion Facilitator
CAX-Cation/Proton Exchanger
ET-Ethylene
GSH-Glutathione
HMs-Heavy Metals
HMAs-Heavy Metal ATPases
HMW-High Molecular Weight
LMWOA-Low-Molecular Weight Organic Acids
MAPK-Mitogen-Activated Protein Kinase
MHX-Magnesium/Proton Exchanger
MTs-Metallothioneins
MTPs-Metal Tolerance Protein
NRAMPs-Natural Resistance Associated Macrophage Protein
RbOHs-Respiratory Burst Oxidase Homologs
ROS-Reactive Oxygen Species
ZIP-Zinc/Iron-Regulated Transporters
Introduction
Heavy Metals (HMs) exists in earth’s crust and sometimes leads to upsurge in upper soil crust due to industrialisation and considered as a major source of toxicity in soil, plant and the whole environment. Metals such as lead (Pb), cadmium (Cd), zinc (Zn), mercury (Hg), arsenic (As), silver (Ag), chromium (Cr), copper (Cu), iron (Fe) and platinum (Pt) have atomic density greater than 4 g/cm3 and are considered heavy metal substances [1]. These metals are major soil pollutants due to their constant non-reactive nature, acute and chronic toxicity. Since these particles don’t degrade or breakdown easily they could affect the biosphere for an extensive period of time. Toxicity and contamination of these metals are one of the major abiotic stresses leading to detrimental effect on plants and ecosystem. Anthropogenic activities such as increased use of sewage discharge, industrial effiuents, superphosphate fertilizers and smelters dust spreading cause heavy metal contamination [2].
The contamination of Heavy Metals (HMs) in soil is one of the key problems that cause damage to plants by changing major physiological and metabolic processes. Notably, few heavy metal ions such as Cu+2, Zn+2, Mn+2, and Fe+2 are essential for plant metabolism at definite concentrations (about 20 ppm), but if in excess it becomes highly toxic [3]. Due to accumulation of HMs, plant characters like seed germination, plant biomass, root length, and chlorophyll biosynthesis seems to deteriorate. Moreover, mineral nutrition, respiration, photosynthetic activity, enzymatic reactions and various other physiological activities are also affected adversely [4]. HMs toxicity in plants leads to increased production of Reactive Oxygen Species (ROS) which disrupts the redox condition of cells resulted in oxidative stress, leading to membrane dismantling, lipid peroxidation, ion leakage, biological macromolecule declination and DNA-strand cleavage [5]. Albeit, the toxic penalty of heavy-metal-induced ROS on plant macromolecules differ and depends on the exposure period, plant growth stage and the concentration of toxic metals in soil. To cope up, plants have developed various survival mechanisms under such abiotic stress conditions.
Phytoremediation is the term generally tells the use of plants and its associated soil microbes to reduce the toxic effects of heavy metals from the soil strata. Phytoremediation consists of phytoextraction, rhizofiltration, phytostabilization, phytovolatization and phytodegradation depending on the mechanisms of remediation. Phytoexraction involves uptake of contaminants by plant roots and subsequent accumulation in the aerial plant parts, followed by harvest and then removal of plant biomass [6]. Rhizofiltration involves the plant roots or seedling for the removal of metals from aqueous wastes. In phytostabilization, plant roots reduce the mobility and bioavailability of metals and organic contaminants in the neighbouring environment preventing their movement in the soil and groundwater or their entry into the food cycle. Phytodegradation consists of use of plants and associated microorganisms to degrade organic pollutants.
Plant response to heavy metal stress involves a convoluted signal transduction system that is activated instantaneously, followed by several signalling networks, such as the Ca-calmodulin pathway, response of phytohormones, ROS mediated signalling, and Mitogen- Activated Protein Kinase (MAPK) mediated phosphorylation cascade. Various reports have claimed that, exogenous application of melatonin leads to enhanced abiotic stress tolerance among various plant species. Whereas, the signalling cascade due to melatonin is still not clear. So, the study of melatonin against HMs needs to be more specified.
Besides this, a novel genetic engineering approaches such as chloroplast gene transformation, gene stacking or gene pyramiding is the next aims to explore. Transgenic plants expressing desirable genes from different organisms are developed to increase the heavy metal remediation efficiency of plants by modifying various targets such as chelators, metal transporters and enzymes. Genetic engineering added quick and considerable changes in the crop improvement by offering a wide collection of novel genes and traits.
Plant Adaptive Mechanisms for Phytoremediation
Uptake and translocation of heavy metals in different plant parts through transporters
Uptake of metals needs metal ions deposition outside roots leads to entry of metal ions in roots and their translocation throughout the shoot through the xylem with mass flow and diffusion method. Several classes of metal transporters play an important role in metal uptake and its homeostasis. It includes Heavy Metal ATPases (HMAs), ZIP (zinc/iron-regulated transporters) family, NRAMPs (Natural resistance associated macrophage protein), CDF (Cation Diffusion Facilitator) and Ca2+. The CPx-type ATPases facilitate the movement of toxic metals like copper (Cu), lead (Pb), and cadmium (Cd) across the plasma membrane by employing energy in the form of ATPP1B- type.
The CDF transporters are involved in mediating the cytoplasmic effiux of transition metal cations such as Zn2+, Cd2+, Co2+, Ni2+ or Mn2+ from the cytoplasm to the endoplasmic reticulum. In the Ca2+/ cation antiporter (CaCA) superfamily, MHX is a vacuolar Mg2+ and Zn2+/H+ exchanger with higher concentrations of MHX protein was present in the leaves of A. halleri than in A. thaliana and was therefore proposed to play a role in Zn vacuolar storage. The plant NRAMP gene family are of particular importance as they’re liable for uptake of the nutritionally vital divalent cations Fe2+, Mn2+, Zn2+, and Cd2+ a toxic metal with no known role in plant growth and development. ABC transporters are ubiquitous transporters involved in a large number of physiological processes. This family is one of the largest protein families with 29, 128, and 48 members in S. cerevisiae, Arabidopsis, and humans, respectively. It is shown that ATP-binding cassette (ABC) transporters may play important roles in Cd tolerance in plants [7]. As major vacuolar phytochelatins (PCs) transporters, ABC transporters (AtABCC1 and AtABCC2) have been proven as well as participate in arsenic (As), mercury (Hg) and Cd resistance [7]. ZIP transporters are referred to as ZRT, IRT-like proteins used in translocation of different ions like Cd, Zn, Fe, Mn depending on the type of substrate and their particular identity. They are involved in the transport of Cd from the soil to roots and its further translocation from the root to the shoot. The expression regulation of both HMA3 and HMA4, reported to transport heavy metals (Cd and Zn) in Cd/Zn hyperaccumulators A. halleri and N. caerulescens [8].
Extracellular Avoidance of Heavy Metals
Avoidance is a basic strategy used to prevent heavy metals in the roots by neutralizing their toxic effects in root cells. Extracellularly, they include roles for
Mycorrhizas
Arbuscular Mycorrhizal Fungi (AMF) which are soil-borne fungi that can widely look up plant nutrient uptake and resistance to several abiotic stress factors. It also helps host plants to grow robustly under stressful circumstances by mediating a sequence of intricate communication events between the plant and the fungus leading to improved photosynthetic rate and other gas exchange-associated traits [9]. Due to their potential to strengthen defence system of the AMF mediated plants, AMF are extensively believed to maintain plant establishment in heavy metal contaminated soils and promote growth and development. Heavy metals can be immobilized in the fungal hyphae that have the capability to fix heavy metals in the cell wall and accumulate them in the vacuole or may chelate with some substances in the cytoplasm and hence decrease metal toxicity in the plants.
Root cell wall exudates
Secretion of root exudates in soil amplifies the external decontamination of the plant under heavy metal stress conditions. Root exudates two types of compounds, (a) high-molecular weight (polysaccharides and proteins) and (b) low-molecular weight (i.e. amino acids, organic acids, sugars and phenolics) compounds; which plays an dominant role in rhizospheric processes. Of which, Low- Molecular Weight Organic Acids (LMWOA) are the most abundant and reactive with metals [10]. An experiment was carried out using three plant species [Poa annua, Medicago polymorpha and Malva sylvestris] for phytoremediation, have been assessed for both metal uptake and excretion of LMWOAs in contaminated environments with different concentrations of Cd, Cu and Zn, M. polymorpha proved to be released the highest concentrations of LMWOAs [10].
Intracellular Deoxification Pathways
When HMs enters the plant tissues, tolerance and detoxification mechanisms are triggered which include metal sequestration and compartmentalization in various intracellular compartments (e.g., vacuole), metal binding to cell membrane, biosynthesis or accumulation of osmolytes and osmoprotectants (e.g., Proline), intracellular complexation or chelation of metal ions by releasing several substances (e.g., phytochelatins (PCs), metallothioneins (MTs) and organic acids) and eventually if all these measures prove futile and plants become overwhelmed with toxicity of Heavy Metal (HM), activation of antioxidant defense mechanisms is pursued [11].
Heavy metal sequestration in vacuoles and trichomes
The plant vacuole is a key storage section for excess heavy metals. HM detoxification mechanism is not only confined to the chelation, but also accumulates and alleviates HMs in the vacuole of cells via formation of High Molecular Weight (HMW) complexes with PCs [6]. Generally, metal ions sequestration is a first line method adopted by organisms to ameliorate toxicity. The immobilized metal ions are bailed out to vacuole for sequestration via transporters, which is mediated by ATP-dependent vacuolar pumps (V-ATPase) as well as via tonoplast transporters [12]. Phytochelatins synthesized from glutathione by phytochelatine sythase, binds cytosolic Cd2+ to form the Low-Molecular-Weight (LMW) complex first, which is moved into the vacuole through a tonoplast-localized ATP-binding-cassette (ABC) transporter. In the vacuole, LMW Cd-complex becomes High- Molecular-Weight (HMW) complex with more Cd2+, which enters into vacuole via direct exchange with protons by tonoplast-localized cation/proton exchanger (CAX) transporters. MTPs (Metal tolerance protein) and NRAMPs existing in the tonoplast mediate passage of metal ions for compartmentation. In case of other chelators like, organic acids and amino acids provides platform for the cytosolic metal concentrations to the safe low metal state (Figure 1).

Figure 1: Vacuolar sequestration of heavy metals in plant cell. [ZIP (zinc/iron-regu- lated transporters), Phytochelatins (PCs), PC synthases (PCS), Low-molecular-weight (LMW), ATP-binding-cassette (ABC) transporter, Cation/proton exchanger (CAX) transporter; Transporters Metal tolerance proteins (MTPs), Natural resistance associ- ated macrophage proteins (NRAMPs), Metallothioneins (MTs)].
Apart from vacuolar sequestration, several reports (Reference) have proved the participation of glandular trichomes in the chelation, sequestration, and detoxification of the metals. Trichomes are specialized unicellular or multicellular structures resultant from the epidermal cell layer, and may have different functions depending on plant species and organs.
Phytochelatins (Pcs) and metallothioneins (Mts)
Higher plants contain two major types of cysteine-rich, metal- binding peptides, the metallothioneins (MTs) and the phytochelatins (PCs). PCs were first identified in yeast as a family of peptides. PCs consist of only three amino acids, glutamine (Glu), cystine (Cys), and glycine (Gly) [13]. The phytochelatins are the foremost widely studied in plants, particularly in reference to Cd tolerance [13]. Phytochelatins are synthesized from glutathione (GSH) through an enzymatic pathway; they chelate heavy metals such as Cd, As, Pb & Hg [14]. PCs play an important role in metal detoxification and in essential metal ion homeostasis in Fe or sulphur (S) metabolism [15]. These are enzymatically synthesized proteins involved in heavy metal detoxification in plants growing in metal-rich soil. Phytochelatins binds to different metals including Cd, Cu, Zn and As through sulfhydryl and carboxyl residues whose biosynthesis is controlled mainly by the glutathione (γ-glutamlcysteinyl-glycine), involves several steps mediated by PC synthase. PC synthase genes were isolated simultaneously by three research groups using different approaches [15]. Similar findings were observed where PCS gene from Populus tomentosa decreased movement of cadmium to the aerial parts of tobacco [5]. PCS-encoding genes mostly of plant origin were used for the transgenesis, resulting in enhanced HM accumulation and tolerance.
MTs are naturally-existing intracellular cysteine-rich major metal-binding proteins, which are used to immobilize, sequesteration, and detoxification of metal ions. The role of plant MTs is generally attributed to the homeostasis of essential metals (Se, Zn, Ni, and Cu), and detoxification of xenobiotic metals (Cd, Hg, and Ag). Introduction of mammalian MT gene into B. compestris, B. napus and N. tobaccum confers Cd tolerance by increasing complexation of metal with MTs and minimizing translocation of Cd to the shoots [13,14]. Besides, it was demonstrated that OsMT2c gene encoding for type 2 MT expressed in the roots, leaf sheathes, and leaves of rice, whereas its weak expression was observed in seeds [16].
Chelating Metals by Organic and Amino Acids and Proline Accumulation
Secretion of organic acid enhances root absorption by the mobilization of heavy metals. Carboxylic acids and amino acids such as citric, malic and histidine are potential ligands for heavy metals and play a role in tolerance and detoxification [17]. Citrate has a high capacity to chelate HM ions and has been well documented in the case of Fe [17]. Genes has been identified related to organic acids exudation and conducted transformation experiments towards improved Al tolerance in plants [18]. Proline, an amino acid, under changing environmental conditions plays different roles such as an osmoprotectant, a protein stabiliser, a metal chelator, an inhibitor of lipid peroxidation and OH• and 1O2 (non radial ROS family) scavenger. Enhanced proline accumulation occurs especially during salt, drought and metal stresses [19]. Thus, proline is not only an essential signalling molecule but also an efficient ROS scavenger.
Antioxidant defense mechanism
Plant cells have developed antioxidant defence mechanism to alleviate the harmful effects of free radicals enclosed several enzymatic antioxidants like superoxide dismutase (SOD), catalase (CAT), ascorbate peroxidase (APX), guaiacol peroxidase (GPX), and glutathione reductase (GR) and non-enzymatic antioxidants like ascorbate (AsA), glutathione reduced (GSH), carotenoids, alkaloids, tocopherols and phenolic compounds (flavonoids, tannins, and lignin) which act as the scavengers. Heavy metal stress upscales the production of ROS and release of free radicals as O -SOD is responsible for dismutation of the two superoxide radicals to H2O2 and O2. While, CAT is an important oxidoreductase enzyme that catalyzes decomposition of H2O2 into H2O and O2 which localized in the peroxisome, involved in detoxifying peroxides generated during photorespiration. When the ROS level exceeds to control the level of antioxidant defence system it leads to oxidative stress (Figure 2). Several researchers have done their experiments regarding ROS scavenging; there was significant rise in SOD, GPX, APX & CAT activities after Pb treatment and their activities were further enhanced with the rising concentration of Pb from 50 to 100 µM [20]. A significant increase in enzymatic (SOD, APX, GR) antioxidant levels was recorded in Sesbania drummondii seedlings, when the seedlings were exposed to Cu, Ni and Zn alone and in combination [21].
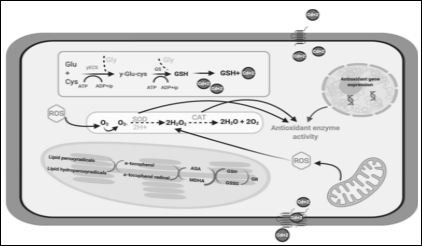
Figure 2: Heavy metal induced oxidative stress, tolerance & detoxification mechanisms in plant cell. [Ascorbic acid (Asa), Catalase (CAT), Cysteine (Cys), C-glutamylcysteinesynthetase (C-ECS), Glutamine(Glu), Glycine(Gly), Glutathione reductase (GR), Glutathione synthetase (GS), Glutathione reduced (GSH), Oxidized glutathione (GSSG), Hydrogen peroxide (H2O2), Monodehydroascorbate (MDHA), Oxygen molecule (O2), superoxide radicals (O2-), Reactive oxygen species (ROS), superoxide dismutase (SOD), heavy metal (A)].
Signal Transduction During Heavy Metal Stress
Signal transduction in plants plays a crucial role to overcome the abiotic and biotic stresses such as salinity, drought, temperature, heavy-metals, pathogens and water stress which possesses detrimental effect on plant growth and development [19]. Being immobile in nature plants cannot move away from the adverse environmental conditions. As a result, plant sense and recognize environmental changes to activate signal transduction in order to initiate molecular and cellular processes of adaptation to the stress. To regulate and complete the signalling pathway, signal transduction mechanism of plants involves.
Signal perception
In recognizing cell, a signal must first be sensed by a receptor to commence transduction. Plant stressors (such as heavy metals) can stimulate signal perception via an external signal (such as calcium), upon which the signals are quickly sent (e.g., through MAP kinases) to the responsible transcription factors [22].
Cellular secondary signal molecules
Phytohormones, Reactive Oxygen Species (ROS), Nitric Oxide (NO), and hydrogen sulfide (H2S) are proposed to be entailed in the response to HM stress and trigger various responses in plants cells e.g. MAP kinases [23]. The calcium ion (Ca2+) as endorsed by different studies acts as a ubiquitous secondary messenger in the normal functioning of plants as well as in response to various environmental stresses [24].
Stress Signal Transduction by Phytohormones
Exogenous application of phytohormones can influence the regulation of the ascorbate-glutathione cycle, transpiration rate, cell division, and the activities of nitrogen metabolism and assimilation, which improve plant growth. Brassinosteroids, ethylene and salicylic acid have been reported to enhance the level of the anti-oxidant system, trim down levels of ROS, lipid peroxidation and improve photosynthesis in plants, when applied exogenously under heavy metal stress [25]. Auxin directly influences plant response to metal stresses by altering auxin homeostasis including auxin stability, transport, and redistribution. Further, it has been reported that PINFORMED1 (PIN1) protein plays an important role in IAA distribution under conditions of HM stress [26]. Enhanced ABA levels were found in response to heavy metals in the leaves of Cd tolerant rice when exposed to Cd [2]. Exogenous application of cytokinins on green algae, Chlorella vulgaris alleviated many deleterious effects of heavy metals (Cd, Pb, and Cu) by reducing metal absorption and stimulating the defense system [27]. The study has shown the contribution of ethylene (ET) in response to abiotic stress, and accelerated production of ethylene in plants exposed to toxic levels of heavy metals [28]. GAs support the development of plant adaptation, resistance to various abiotic stresses as well as reported as iron (Fe) translocation inhibition by suppressing OsYSL2 gene expression in addition to regulation of Fe transport and translocation [29]. Salicylic acid was considered to reduce the ROS level and lipid peroxidation, whereas it increased the electrolyte leakage, chlorophyll content, total lipids and linolenic acid contents [30].
Of all, a master regulator - melatonin (N-acetyl-5- methoxytryptamine) not only orchestrates numerous physiological activities in plants but also acts in defence responses against biotic and abiotic stresses [31]. In plants, it shares the same initial biosynthesis compound with auxin, and therefore functions as indole-3-acetic acid like hormones [32]. Melatonin is also considered as a plant growth regulator because of its specific physiological functions such as regulation of the growth of roots, shoots, explants, seed germination, rhizogenesis and delay in leaf senescence [33, 34]. During abiotic stress conditions, it has been revealed that melatonin deploys by plant as an effective free radical scavenger against harmful Reactive Oxygen (ROS) and Reactive Nitrogen (RNS) Species [35]. Melatonin has been proposed as a new plant hormone since its receptor (PMTR1) has been identified in Arabidopsis. This receptor, located in the plasma membrane, is able to interact with G-protein α subunits, activating Respiratory burst oxidase homologs (RbOHs) and promoting Ca2+ and K+ fluxes that result in stomatal closure [34]. Heavy metal- stressed plants regulate the expression of melatonin synthesis-related genes, such as OsASMT, to enhance heavy metal tolerance as well as increased M. truncatula growth under Pb stress [36]. Researchers have reported effects of Melatonin on heavy-metal-treated plants as per following (Table 1).
Genetic Engineering to Enhance Phytoremediation in Plants
Genetic engineering of plants for phytoremediation involves various metal transporters, chelators and enzymes. It has been reported that the majority of membrane transporter families aids the nutrient metal uptake, transport, and homeostasis in plants [37-39]. Transgenic plants can be developed by inserting transgenes into any of the three genomes of plants, for example, chloroplast, nuclear, plastidal or mitochondrial. To improve phytoremediation considerably,

Table 1: Interrelation between melatonin and heavy metal treatment in plants.
Gene stacking
The strategy refers to the process of combining two or more gene of interest in the genome of a single plant. It has been reported that dual-transgenic A. thaliana plants overexpressing both Phytochelatins (PCS derived from garlic) and γ-g lutamylcysteine synthetase (γ-ECS from yeasts) led to increased PC levels and exhibited significantly higher Cd/As tolerance and accumulation than single transgenic and non-transgenic controls [40],
Transformation of chloroplast genome
This approach was used first time to improve the capacity of plants for phytoremediation, achieved by combining two genes merA (which code for mercuric ion reductase) and merB (organomercurial lyase) into the chloroplast genome of Nicotiana tabacum [41]. Stable integration of the merAB operon into the chloroplast genome triggered to enhanced tolerance up to 500 µM phenylmercuric acetate (PMA) [42]. It has been reported that overexpression of At γ-tocopherol methyltransferase gene (γ-TMT) in transformed tobacco chloroplasts resulted in a higher concentration of α-tocopherol accumulation in TMT plants than in the wild-type plants under Cu stress. As a consequence, transplastomic plants grew better and exhibited increased tolerance related to antioxidant defense [43].
Genetically Modified Plants and Phytoremediation
Genetic engineering is a prevailing means that may quicken the evolution of new plant lines with characters required in phytoremediation. Transgenic plants expressing various genes from different sources have been developed to increase heavy metal tolerance, accumulation, and volatilization that facilitate more effective heavy metal phytoremediation (Table 2). The main classes of metal chelators include metallothioneins (MT), phytochelatins (PCs), which are peptides enzymatically synthesized from glutathione, and Low Molecular Organic and Amino acids (LMWOA). Only MTs are direct products of gene expression among them. During external HM stress, Metallothionein gene IlMt2a from halophyte Iris lactea var. chinensis was integrated into the Arabidopsis genome which triggered greater tolerance to Cd (up to 100 µM) and Cu (50 µM and 100 µM) [50,51].

Table 2: Transgenic plants developed for heavy metal phytoremediation.
PCs are low molecular weight cysteine-rich metal-binding peptides which is synthesised from Glutathione. Under Abiotic stress condition, glutathione (GSH) acts as an antioxidant and regulatory molecule in plants. GSH plays a central role in metal chelation (by itself or as a precursor for PCs), as well as through its antioxidant capacities. Overexpression of LmgshF gene under CaMV35S promoter in Oryza sativa L. showed tolerance up to 6.07 fold at 200 mM NaCl treatment [52]. The melatonin-rich transgenic Arabidopsis plants overexpressing alfalfa SNAT (a melatonin synthetic gene) showed more tolerance than wild-type plants under Cd conditions [53]. Expression of MsASMT1 (N-acetylserotonin methyltransferase) gene into alfalfa, which considerably increased the endogenous melatonin content [54].
Conclusion and Futrher Perspective
Remarkable development has been made in identifying and comprehensing the role of major components like metal transporters, metal chelators, mycorrhizae, signaling molecules and phytohormones that ensure heavy metal tolerance to plants. To avoid health hazards, existing remediation alternatives focus on minimizing the concentration of heavy metals in soil and the food chain. There has been an important role of melatonin emerged from the numerous studies to mobilize toxic metals through phytochelatins, their transport and sequestration resulting in a high degree of plant tolerance against heavy metal stress. However, further studies are needed to provide genetic evidence in support of involvement of melatonin to elucidate the subsequent signalling cascades in plants. Plant-engineering strategies for heavy metal tolerance depend on the expression of gene(s) whose product(s) are involved either in signalling and regulatory pathways or in the synthesis of functional and structural proteins and metabolites that confer HM stress tolerance. This review has focused on recent proofs that specify potential cellular and molecular methods that may be involved in the resistance and tolerance of plants under heavy metals stress. An apprehension of HM tolerance mechanism in plants is being refined by the growing interest in molecular genetics as well as transgenic plants have exhibited enhanced HM tolerance. Modification in different characteristics in plants such as metal uptake, transport, signalling, accumulation and plant’s tolerance to metals through genetic engineering imparts new possibilities of phytoremediation.
There is a need of efforts from plant physiologists, agronomists, soil scientists, molecular biologists, microbiologists, environmental scientist and government regulatory to achieve further improvements in phytoremediation and to reduce load of abiotic stresses in the ecosystem.
References
- Onakpa MM, Njan AA, Kalu OC (2018) A Review of Heavy Metal Con- tamination of Food Crops in Nigeria. Ann Glob Health 84: 488-494.
- Kim YH, Khan AL, Kim DH, Lee SY, Kim KM (2014) Silicon mitigates heavy metal stress by regulating P-type heavy metal ATPases, Oryza- sativa low silicon genes, and endogenous phytohormones. BMC Plant Biol 14: 1-13.
- Singh S, Parihar P, Singh R, Singh VP, Prasad SM (2016) Heavy Metal Tolerance in Plants: Role of Transcriptomics, Proteomics, Metabolom- ics, and Ionomics. Front Plant Sci 6: 1143.
- Pourrut B, Perchet G, Silvestre J, Cecchi M, Guiresse M (2008) Potential role of NADPH-oxidase in early steps of lead-induced oxidative burst in Viciafaba roots. J Plant Physiol 165: 571-579.
- Chen Y, Liu Y, Ding Y, Wang X, Xu J (2015) Overexpression of PtPCS enhances cadmium tolerance and cadmium accumulation in tobacco. Plant Cell Tissue Organ Cult 121: 389-396.
- Jabeen R, Ahmad A, Iqbal M (2009) Phytoremediation of Heavy Metals: Physiological and Molecular Mechanisms. Bot Re 75: 339-364.
- Park J, Song WY, Ko D, Eom Y, Hansen TH (2012) The phytochelatin transporters AtABCC1 and AtABCC2 mediate tolerance to cadmium and Plant J 69: 278-288.
- Mishra S, Mishra A, Kupper H (2017) Protein Biochemistry and Expres- sion Regulation of Cadmium/Zinc Pumping ATPases in the Hyperac- cumulator Plants Arabidopsis halleri and Noccaeacaerulescens. Front Plant Sci 8: 1-13.
- Birhane E, Sterck F, Fetene M, Bongers F, Kuyper T (2012) Arbuscu- lar mycorrhizal fungi enhance photosynthesis, water use efficiency, and growth of frankincense seedlings under pulsed water availability condi- Oecologia 169: 895-904.
- Montiel-Rozas MM, Madejon E, Madejon P (2016) Effect of heavy metals and organic matter on root exudates (low molecular weight organic ac- ids) of herbaceous species: An assessment in sand and soil conditions under different levels of Environ Pollution 216: 273-281.
- Manara A (2012) Plant responses to heavy metal toxicity in Plants and Heavy In: Springer briefs in molecular science. Furini A, editors. Springer, Dordrecht, Netherlands 2753.
- Sharma SS, Dietz KJ, Mimura T (2016) Vacuolar compartmentalization as indispensable component of heavy metal detoxification in plants. Plant Cell Environ 39: 1112-1126.
- Cobbett CS (2000) Phytochelatins and their roles in heavy metal detox- Plant Physiol 123: 825-832.
- Hall JL (2002) Cellular mechanisms for heavy metal detoxification and J Exp Bot 53: 1-11.
- Cobbett C, Goldsbrough P (2002) Phytochelatins and metallothioneins: Roles in heavy metal detoxification and homeostasis. Annu Rev Plant Biol 53: 159-182.
- Liu J, Shi X, Qiana M, Zhenga LC, Yan X (2015) Copper-induced hy- drogen peroxide upregulation of a metallothionein gene, OsMT2c, from Oryza sativa confers copper tolerance in Arabidopsis thaliana. J Haz- ard Mater 294: 99-108.
- Clemens S (2001) Molecular mechanisms of plant metal tolerance and Planta 212: 475-486.
- Zhou G, Pereira JF, Delhaize E, Zhou M, Magalhaes J (2014) Enhancing the aluminum tolerance of barley by expressing the citrate transporter genes SbMATE and FRD3. J Exp Bot 65: 2381-2390.
- Hakeem KR, Rehman RU, Tahir I (2014) Plant signaling: Understanding the molecular crosstalk. (Eds.). Springer India.
- Bharwana SA, Ali S, Farooq MA, Iqbal N, Abbas F (2013) Alleviation of lead toxicity by silicon is related to elevated photosynthesis, antioxi- dant enzymes suppressed lead uptake and oxidative stress in cotton. J Bioremed Biodeg 4: 4.
- Israr M, Jewell A, Kumar D, Sahi SV (2011) Interactive effects of lead, copper, nickel and zinc on growth, metal uptake and antioxidative me- tabolism of Sesbaniadrummondii. J Hazard Mater 186: 1520-1526.
- Sinha AK, Jaggi M, Raghuram B, Tuteja N (2011) Mitogen-activated pro- tein kinase signaling in plants under abiotic stress. Plant Signal Behav 6: 196-203.
- Luo ZB, He J, Polle A, Rennenberg H (2016) Heavy metal accumula- tion and signal transduction in herbaceous and woody plants: Paving the way for enhancing phytoremediation efficiency. Biotechnol Adv 34: 1131-1148.
- Jalmi SK, Bhagat PK, Verma D, Noryang S, Tayyeba S (2018) Travers- ing the Links between Heavy Metal Stress and Plant Signaling. Front Plant Sci 9: 12.
- Verma V, Ravindran P, Kumar PP (2016) Plant hormone-mediated regu- lation of stress responses. BMC Plant Biol 16: 86.
- Li K, Kamiya T, Fujiwara T (2015) Differential roles of PIN1 and PIN2 in root meristem maintenance under low-B conditions in Arabidopsis thali- ana. Plant Cell Physiol 56: 1205-1214.
- Piotrowska-Niczyporuk A, Bajguz A, Zambrzycka E, Godlewska ZB (2012) Phytohormones as regulators of HM biosorption and toxicity in green alga Chlorella vulgaris (Chlorophyceae). Plant Physiol Biochem 52: 52-65.
- Khan MIR, Nazir F, Asgher M, Per TS, Khan NA (2015) Selenium and sulfur influence ethylene formation and alleviate cadmium-induced oxi- dative stress by improving proline and glutathione production in wheat. J Plant Physiol 17: 9-18.
- Wang B, Wei H, Xue Z, Zhang WH (2017) Gibberellins regulate iron de- ficiency-response by influencing iron transport and translocation in rice seedlings (Oryza sativa). Ann Bot 119: 945-956.
- Kazemi N, Khavari-Nejad RA, Fahimi H, Saadatmand S, Nejad-Sattari T (2010) Effects of exogenous salicylic acid and nitric oxide on lipid perox- idation and anti-oxidant enzyme activities in leaves of Brassica napus under nickel stress. Sci Hortic 126: 402-407.
- Arnao BM, Hernandez-Ruiz J (2019) Role of Melatonin to Enhance Phy- toremediation Capacity. Appl Sci 9: 5293.
- Fan J, Xie Y, Zhang Z, Chen L (2018) Melatonin: A multifunctional factor in plants. Int J Mol 19: 1528.
- Arnao BM, Hernandez-Ruiz J (2018) Melatonin and its relationship to plant hormones. Ann Bot 121: 195-207.
- Arnao MB, Hernández-Ruiz J (2019) Melatonin and reactive oxygen and nitrogen species: A model for the plant redox Melatonin Res 2: 152-168.
- Tan DX, Manchester LC, Reiter RJ, Qi WB, Karbownik M (2000) Sig- nificance of melatonin in antioxidative defense system: Reactions and Biol Signals Recept 9: 137-159.
- Zhang X, Zhang H, Zhang H, Tang M (2020) Exogenous Melatonin Ap- plication Enhances Rhizophagus irregularis Symbiosis and Induces the Antioxidant Response of Medicago truncatula Under Lead Front Microbiol 11: 516.
- Nawaz MA, Jiao Y, Chen C, Shireen F, Zheng Z (2018) Melatonin pre- treatment improves vanadium stress tolerance of watermelon seedlings by reducing vanadium concentration in the leaves and regulating mel- atonin biosynthesis and antioxidant related gene expression. J Plant Physiol 220: 115-127.
- Hasan MK, Ahmmad GJ, Yin L, Shi K, Xia X (2015) Melatonin mitigates cadmium phytotoxicity through modulation of phytochelatins biosynthe- sis, vacuolar sequestration, and antioxidant potential in Solanum lycop- ersicum L. Front Plant Sci 6: 601.
- Paulose B, Jaiwal P, Dhankhe OP (2009) Heavy Metal Transporters in Plants. Plant membrane and vacuolar transporters, CABI, Wallingford/ Cambridge. Pg no: 213-238.
- Guo J, Dai X, Xu W, Ma M (2008) Overexpressing GSH1 and AsPCS1 simultaneously increases the tolerance and accumulation of cadmium and arsenic in Arabidopsis thaliana. Chemosphere 72: 1020-1026.
- Ruiz ON, Hussein SH, Terry N, Daniell H (2003) Phytoremediation of Or- ganomercurial Compounds via Chloroplast Genetic J Plant Physiol 132: 1344-1352.
- Hussein HS, Ruiz ON, Terry N, Daniell H (2007) Phytoremediation of mercury and organomercurials in chloroplast transgenic plants: En- hanced root uptake, translocation to shoots, and volatilization. Environ Sci Technol 41: 8439-8446.
- Jin S, Daniell H (2014) Expression of γ-tocopherol methyl transferase in chloroplasts results in massive proliferation of the inner envelope mem- brane and decreases susceptibility to salt and metal-induced oxidative stresses by reducing reactive oxygen species. Plant Biotechnol J 12: 1274-1285.
- Das N, Bhattacharya S, Maiti MK (2016) Enhanced cadmium accumula- tion and tolerance in transgenic tobacco overexpressing rice metal tol- erance protein gene OsMTP1 is promising for phytoremediation. Plant Physiol Biochem 105: 297-309.
- Zhang J, Zhang M, Tian S, Lu L, Shohag MJI (2014) Metallothionein 2 (SaMT2) from Sedum alfredii Hance confers increased Cd tolerance and accumulation in yeast and tobacco. PloS one 9: e102750.
- Shukla D, Kesari R, Tiwari M, Dwivedi S, Tripathi RD (2013) Expres- sion of Ceratophyllum demersum phytochelatin synthase, CdPCS1, in Escherichia coli and Arabidopsis enhances heavy metals Protoplasma 250: 1263-1272.
- Kiyono M, Oka Y, Sone Y, Tanaka M, Nakamura R (2012) Expression of the bacterial heavy metal transporter MerC fused with a plant SNARE, SYP121, in Arabidopsis thaliana increases cadmium accumulation and Planta 235: 841-850.
- Yang JL, Wang YC, Liu GF, Yang CP, Li CH (2011) Tamarix hispida metallothionein-like ThMT3, a reactive oxygen species scavenger, in- creases tolerance against Cd2+, Zn2+, Cu2+, and NaCl in transgenic Mol Biol Rep 38: 1567-1574.
- Lang M, Hao M, Fan Q, Wang W, Mo S (2011) Functional characteri- zation of BjCET3 and BjCET4, two new cation-efflux transporters from Brassica juncea L. J Exp Bot 62: 4467-4480.
- Gu CS, Liu LQ, Zhao YH, Deng YM, Zhu XD (2014) Overexpression of Iris. lactea var. chinensis metallothionein llMT2a enhances cadmium tolerance in Arabidopsis thaliana. Ecotoxicol Environ Saf 105: 22-28.
- Gu CS, Liu LQ, Deng YM, Zhu XD, Huang SZ (2015) The heterologous expression of the Iris lactea chinensis type 2 metallothionein IlMT2b gene enhances copper tolerance in Arabidopsis thaliana. Bull Environ Contam Toxicol 94: 247-253.
- Srivashtav V, Jha S, Parekh V (2019) Overexpression of LmgshF from Listeria monocytogenes in Indica Rice Confers Salt Stress Tolerance. Russ J Plant Physiol 66: 911-921.
- Gu Q, Chen Z, Yu X, Cui W, Pan J (2017) Melatonin confers plant tol- erance against cadmium stress via the decrease of cadmium accumu- lation and reestablishment of micro RNA-mediated redox homeostasis. Plant Sci 261: 28-37.
- Cen H, Wang T, Liu H, Wang H, Tian D (2020) Overexpression of MsASMT1 Promotes Plant Growth and Decreases Flavonoids Bio- synthesis in Transgenic Alfalfa (Medicago sativa L). Front Plant Sci 11: 489.
Citation: Mehta V, Kansara R, Srivashtav V, Savaliya P (2021) A Novel Insight into Phytoremediation of Heavy Metals through Genetic Engineering and Phytohormones. J Nanosci Nanomed Nanobio 4: 010.
Copyright: © 2021 Mehta V, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and re- production in any medium, provided the original author and source are credited.
